
Understanding the Mind
Preface
Understanding the MIND points-out the divergence between fixed brain functions and volitional actions that are directed by the conscious MIND. We commonly do not think about how to carryout actions unless we are conditioning new movement patterns. For example, the golfer relies on trial and error and a coach. This presentation demonstrates how components of the MIND emerge from brain function to produce consciousness, emotion, cognition, decision making and the content of speech. The conscious MIND designates action targets as goals and initiates the motor action while the BRAIN generates the pattern of movements via reflexes and hardwired patterned movement generators from basal ganglia and cerebellum. The role of the brain is to maintain homeostatic control of body functions, provide mobility and control movement patterns that serve as a platform for the emergence of MIND that is expressed in behaviors. The most basic MIND function is the generation of CONSCIOUSNESS. Most sensory systems generate inputs to the brain that lead to emotion-related memories. The ultimate capacity of the MIND is in decision-making for defining attention to specific sensory modalities and in initiating willful behaviors that include generation of speech content. The integration of emotional memories into cognition yields understanding that generates intellect and perception. MIND-directed values and actions are represented in behavior of each individual during life and are held in the MIND of others as the individual's SOUL. Action-values, that are defined by the MIND in life, engender a SPIRIT that inspires others into action for some time after the brain ceases to function. Refer to STUDY MATERIALS (65 FREE modules) for detailed descriptions of how neurons function and incorporate their actions into circuitry generating MIND actions through neurons. Dean E. Hillman, PhD See also howthebrainworks.net |
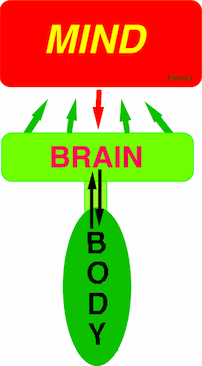
MIND Brain and Body |
WHAT IS THE MIND?The MIND is a distinct entity from the brain itself. Brain circuitry serves as the foundation for homeostasis and behaviors as well as the emergence of the MIND from the Brain. The MIND is a "consciousness producing energy-state" of the cerebral cortex resulting from synchronous relaying of sensory inputs to pyramidal neurons of sensory cortices. This energy-state yields self-awareness out of sensory stimuli or from memories. Accentuation of sensory memories occurs through the correlation of emotions to sensory inputs or behavioral events. Emotions are the one factor that stimulates memories bringing the most visible events to the center of attention. The hippocampus is the emotion determination center by comparing inputs of facial expressions, body postures, site locations or sounds to incoming sonsory experiences. The hippocampal pathway sends out signals to the amygdala, dorsal thalamus and hypothalamus, all eliciting emotional responses in their target areas. This emotion generating part of the MIND defines values and associates sensory inputs to stored emotional memories. The MIND classifies sensory events as friend or foe or as promoting pleasure or displeasure. The hypothalamus activates the autonomic nervous system in the role to sustain homeostasis. The dorsal thalamus signals conscious emotions through connections to the neocortex (the cingulate gyrus) where emotional feelings get stored. The amygdala innervates the prefrontal cortex (cognition) and insula (vision) controlling behaviors. Sequential cascades of sensory signals onto groups of cerebral pyramidal cells generate inputs for cognition and behavioral decision-making. It is from these cascades that the intellect of MIND arises and decisions are made for initiating volitional behaviors and expression of speech content. The differentation between the MIND and brain function is obvious in speech content versus speech ability. Functional processing by the brain is based in circuits that are fixed genetically or by developmetal conditioning. Learning to speak is a brain conditioned function. Composing and giving a speech is from the MIND. Another factor of the MIND is volition versus producing fixed motor patterns. The MIND can intentionally direct individual movements in a process of learning through conditioning complex reflexes and volitionally re-programming behaviors. MIND operations emerge as volitional actions in contrast to hardwired-reflexes of spinal cord and brainstem or to conditioned reflexes that are stored as programmed patterned movements that we recognize as individual behavioral patterns and the ability to speak. Nevertheless, the MIND selects and initiates target goals for behavior but does not define the motor patterns of movement except through motor learning processes. Essentially, the MIND conditions programmed control through learning processes that utilize conscious trials and error evaluations. By directing sensory attention to movement order (most effective when guided by a coach), the MIND evaluates movement consequence and uses these results to enhance movement patterns as motor-learning. Once the movement pattern is learned it becomes a conditioned reflex of automatic programmed movements and is not directed by MIND, although the Mind initializes pattern movements. Thus, conditioned patterned movements require stringent MIND-conscious attention in order to suppress expression. (See Source of MIND at end of this section and in MATERIALS). The activated MIND yields:
These components form the basis of INTELLECT and expression of PERSONALITY D E Hillman 5/1/ 2008; Edited 2015 See also howthebrainworks.net
|
|
MIND & CONSCIOUSNESSThe mark of MIND is consciousness. The common base for recognizing the existence of MIND is the awareness of sensory stimuli and this are usually followed by voluntary-motor activity. Consciousness occurs in sensory cerebral cortices as self-awareness when the dorsal thalamus gates packets of sensory input to cerebral cortical pyramidal cells. Consciousness begins to emerge when the dorsal thalamus drives sensory inputs to pyramidal cells of the primary cerebral cortices at over 5 times a second. Awaking of the MIND to consciousness occurs when dorsal thalamic neurons send sensory input to cortical pyramidal neurons at 15 to 40 times a second. Consciousness arises from pyramidal cell activity occurring every 30 to 70 milliseconds of data in bursts. The dorsal thalamus acts by timing of relay-inputs to respective sensory modalities. The two main sensory sources, activating consciousness, are the external environment and from within ones self. Vision, audition, taste and olfaction, report on the external world while sensory-self inputs arise from body surfaces and internal organ receptors. Skin, muscle, tendon and bone are the major contributors to bodily sensory activations reaching the thalamus with relay to the cerebral cortex. Sound, pain and physical movement are the most responsive generators of consciousness from sleep or unconscious states. In parallel, the MIND initiates automatic motor control for eye movements and extensor muscles (e.g. standing). Hyper-stimulation of any one sensory system focuses MIND-awareness to that system. Nevertheless, the MIND can direct focus to other sensory inputs even though the signals maybe weak. Most often, our senses operate at subliminal levels of AWARENESS providing minimal MIND determined attention. Without prominent sensory inputs, the MIND recalls memories and these act to sustain consciousness.
|
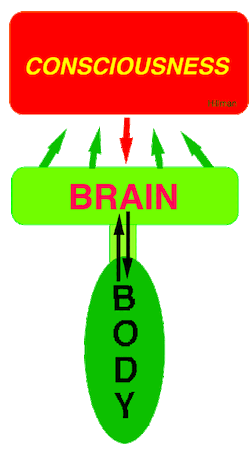
Consciousness Emerges out of Sensory Brain Function. 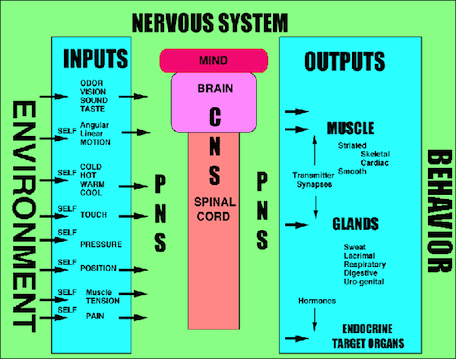
Sensory input and CNS output. |
MIND & MEMORYAlternatively, the conscious MIND recalls memories of previous sensory experiences and compares them to currently received inputs providing reinforcement to consciousness. MEMORY is the central pillar of the conscious MIND. Memory is the grist for planning and imagination by the MIND. The MIND has the ability to assemble, store and recall MEMORY from primary and secondary sensory cortices. The conscious and dreaming MIND can also contributes to memory storage. Sensory inputs are processed in primary cortices for ongoing differences between memories of current sensory events to previous inputs. Sensory modalities target different regions of the cerebral cortex suggesting that memory is a distributed process and is associated to multiple parts of the cortex. Without inputs from sensory systems or memory recall, the MIND has difficulty in sustaining consciousness and lacks ability to make decisions. In addition, associational cortices surround the primary sensory cortices and process details of each respective sensory modality. An example is vision where area 17 receives the primary signal but the adjacent areas 18 and 19 represent color and image detail. MEMORY STORAGEThe basis of memory is in mechanisms for memory storage and retrieval. MEMORY formation has five fundamental domains: 1) increased synaptic activity in pathways ending as NMDA synapses, 2) functional activity in these circuits produce long term potentiation (LTP) that is related to removal of a magnesium ion from the NMDA receptor channel allowing calcium to enter the cytosol, 3) this cytosolic calcium acts as an intermediate signal to early gene expression (c-fos, c-fyn, etc), 4) these signals produce specific synaptic proteins that are required for learning by changing synapses and 5) a physical reorganization of the size of synaptic sites on each neuron. This synaptic size adjustment strengthens or diminishes the inputs between same sources to establish fixed data streams that are related to each memory event. Thus, the basis of memory inclusion appears to be a shift in the relative size of synaptic receptor sites on the same neuron. Because the total number of receptor molecules on each neuronal type appears to remain constant (Hillman & Chen), the translation of learning into memory appears to be a shift in the relative sizes of receptor sites to the same neuron while the afferent axons can markedly increase their projection target contact site. Shifting of input strength of specific synaptic sites alters activity across different components of neuronal circuitry representing inputs such as each sensory modality. Each of these sensory modalities is realized consciously at the projection target site in the cerebral cortex. Visual memories, especially, reveal the distributed process of memory storage. Images are stored as collage-scenes of multiple small images or shapes that are literally drawn-out by eye movements. The foveal retina follows the margins of faces or objects combining vision and eye movement into scenes. The relatively small area of the fovea, as compared to reflex-inducing peripheral retina, restricts formation of scene images (although some report photographic memory). Therefore, scenes are compiled from small area images with the details being left out. An example is reading where images are words and phrases. Panoramic views of each page are generated by memories for eye and head movements allowing small image areas to be linked through motor actions. Rapid eye movements (REM) during sleep may represent the consolidation of image memories. Storage of scene imagery appear to be formed by coupling motor actions of eye movements with small detailed images. Being that images are not being formed during sleep, they do not activate the MIND but allow visual memories to be written into circuit activity. Auditory memories are composed of sound frequency patterns that are formed as sound durations and intervals, variations in loudness and differences in direction. Sound is very strong in alerting the MIND. Olfaction does not pass through the thalamus but has its own target direct to the olfactory cortex and olfactory bulb. Smells are non-alerting to the MIND but act secondarily through irritations they produce. Fire victims result because the MIND fails to be activated primarily for smoke but only secondarily by irritation. Taste activation of the mind is largely reflexive to the vomiting motor control center and a small area of the postcentral gyrus near the lateral fissure. Touch represents a broad number of sensory types and locations with input gating to the postcentral gyrus of the cerebral cortex. Receptors on the body surface are represented in the skin with deep receptors occupying muscle, joints and tendons. MEMORY RETRIEVALThe most retrievable memories are associated with high-emotional levels as are those that are most lasting. Memory events are associated in time and location by linkage to hippocampal stored images that are specific to emotions. Memories are retrieved through the sensory-cortex by sending outputs through the hippocampus. Most memories are lost or distanced if they are not reinforced by associational recall or lack an emotional counterpart. The association of events into cognitive frameworks of understanding provides the most meaningful memories for retrieval of understanding. MEMORY & FORGETTINGMemory of events is continually undergoing a natural progression of forgetting. Parking your car each day in a large garage of scattered stalls produces numerous memories of locations. We recall the most recent memory of parking by the strength of the memory for locations among previous days. The forgetting process is the clearing of “attention memory” by a temporal process of declining strength for memory recall of previous image memories. Nevertheless, memories associated with strong emotional events remain most retrievable in recall. Also, the more often that an event is recalled, the stronger is the stability of it’s memory. Repetitive recalling of memory is a common technique for learning and reinforcing long-term memory. |
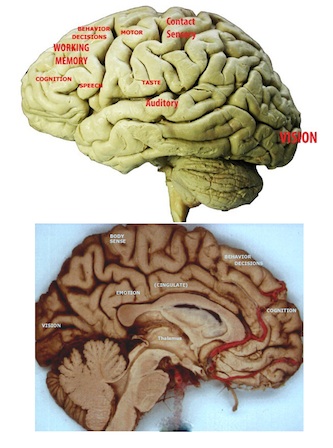 > >Neocortical sensory regions and thalamic targets. |
MIND & EMOTIONBesides the storing of sensory memories, the MIND associates these inputs with emotions that are coincident with memory events. Sensory images of facial expressions, body postures, site locations as well as sound are stored as templates in the hippocampus. These are used in comparisons to incoming facial images, places and sounds. Visual and auditory cortices connect directly to the hippocampus through the occipital-temporal fasciculus (inferior longitudinal fasciculus) that extends from the visual cortex to the medial temporal lobe passing under the parahippocampal gyrus. This input projects to the hippocampus through the perforant path from the entorhinal cortex. (See Vision and emotion in Materials for details) Visual and auditory inputs are incorporated into emotion-memory by the hippocampus with the amygdala determining involuntary behavioral responses through the hypothalamus. Hypothalamic projections provide control to autonomic motor neurons of the brainstem for parasympathetic and for sympathetic control via the spinal cord (lateral horn). A second output of the hippocampus is to the amygdala that projects to the frontal cortex, prefrontal cortex and insula. Since this pathway does not go through the dorsal thalamus, it may be a modulator of cortical motor inputs rather than moving sensory information through the dorsal thalamus. Thus, the amygdala up-regulates premotor regions for aggressive actions. Learning about these transmitters and receptors of the amygdalar/cortical connection will be important for understanding behavioral actions related to societal survival by averting violence that plagues mankind. The third output represents MIND-Emotion. This output is from the hippocampus and projects via the fornix to the hypothalamic mammillary bodies. From here, the projection is relayed by the dorsal thalamic anterior group (AG) thalamic nuclei. This pathway gates emotions to the cingulate gyrus as the temporal synchronizing center for emotion-inputs. The cingulate gyrus is a ready recall source of emotional events and images for use in cognition and motor decision-making. Notably, this pathway appears to be the essential pathway for “remembering”. |
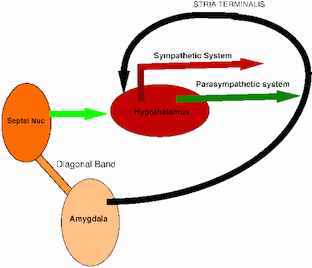
Limbic Control of the Autonomic Nervous System. 
Direct Connection of Amygdala to Cortex 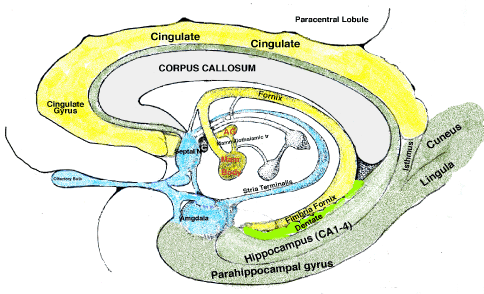
Limbic Components. |
MIND & VALUESSensory-motor reflex organization of the BRAIN lacks a value system; yet, a MIND-VALUE system is developmentally engendered and becomes the foundation of the conscious Mind. The MIND adopts VALUES establishing the character of the individual’s MIND. What is pleasure to one individual may not be to another. VALUES are recalled in association with each memory according to our inherently gained ethical standards. VALUES are influenced by culture as well as by maternal/paternal inputs that instill basic values “for the greater good of the species”. Values arise from positive as well as negative reinforcements. Values are determined by pleasure or displeasure principles. Society and governments largely uses shaming to drive values, negatively, to individual MINDs. Parental guidance varies from highly negative to positive reinforcement. Negative values are often instilled as physical punishments as well as verbal insults of the individual's character. The MIND associates motor outcomes to memories and uses them to establish ACTION-VALUES. Outcomes of motor decision as well as associated reflex actions and programmed movements are stored in emotional memory according to their value level. The MIND uses these VALUES to make decisions for initiation of movements that are expressed in our behavior and speech. Emotions continue to develop over time as changes in levels of neurochemical transmitters and development of their receptors leading to habits and addictions. Thus, the MIND can easily looses control to conditioned reflexes resulting in chemical addictions. MIND & MOTIVATIONMIND-determined motivations arise from three principal sources: need for food, sexual fulfillment, and self-esteem. Willful behaviors are based on each individual’s need to fulfill the three basic needs. The principal drive of the MIND is self-esteem and this underlies all willful behaviors. From birth, the MIND is seeking to fulfill the need for food. As the conscious MIND matures, the reinforcement of self-esteem is recognition from others. The individual’s needs for reinforcing self-esteem repeat every few minutes to hours. The child seeks its’ mother less often with age; adults rely on a pet, partner, child, work associates or themselves as self esteem providers. Curiously, coffee breaks occur at about 1 1/2 to 2 hours. Fulfillment of the self-esteem need comes from within self, but is reinforced externally from others. These reinforcements can be positive, but too often are negative, and as a result are the only means for the individual to gain self-esteem. Whether these individuals become bullies and position themselves outside of nominal society or else subordinate themselves, depends on the strength of negative assaults and the strength of the individual’s personality. The direction of personality development by positive reinforcement is most successful but must be done with an understanding of the stimulus and the consequence of influences on behavior. |
|
MIND AND COGNITIONThe INTELLECT of mankind is in cognitive capacity. The MIND has resources to assemble unqualified sensory and motor memories into "frameworks of understanding". The MIND recalls memories according to emotions, spatial-temporal domains and consequences of actions, and then uses these in cognition to conceptualize temporal processes and structural order. There are several cortical regions involved in understanding. The longest recognized decoding of auditory and visual language is Wernickes area that lies at the confluence of various sensory input regions that are often designated as the angular gyrus, area 39. This region brings together all of the sensory inputs (decoding region) into understanding. A second cognition region is working memory in the frontal lobe as region 46. This region is believed to be an active site bringing together cognitive capacity for expression through adjacent Broca speech region for encoding understanding. The most prominent cognitive region for cognition is defined by the medial dorsal thalamic (MD) relay nuclear gate to the prefrontal cortex in the frontal lobe. Memories of emotional content are processed for COGNITION in passage to the medial dorsal thalamus (MD). This relay nucleus gates inputs, temporally, to pyramidal cells of the prefrontal region of the neocortex. Pyramidal cells are resynchronized by a presumed voltage-gated calcium channel activation. The reason for this cyclic massive depolarization is hypothesized to prevent residual pyramidal cell depolarization of the previous thalamo-cortical relay and allow rapid shifts of the MIND between different cognitive thoughts. The actual stored information is held in structural connectivity while the physiological component acts as a freely adaptable processor that includes long term potentiation (LTP) for temporary holding of new information before it is encoded into physical memory. The cascading of memory and emotion to the cognitive brain integrates understanding of order for processes and structures by expanding thoughts back into memories. The cognitive cascade yields a base for the power to reason, to formulate cognition and to make decisions. The frontal lobe (prefrontal cortex) extrapolates sensory memories, motor experiences and emotional responses into PREDICTIONS yielding higher levels of INTELLECT. The intellect is reinforced by sensory inputs that are driven to foremost attention by emotion. Foretelling consequence as PREDICTION is the mark of the intellectual MIND. Fundamental to this process is the ability to IMAGINE by projecting-understanding into a VISION that extends beyond the known. |
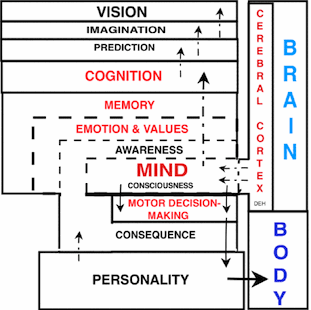
COGNITION Leads to Imagination and Vision. 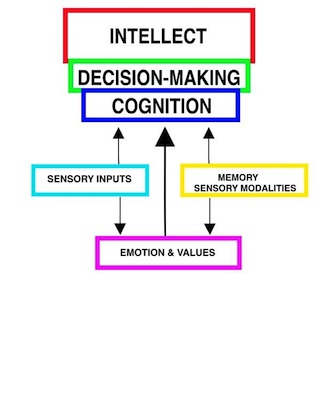
Intellect Results from COGNITION. |
MIND AND DECISION-MAKINGDecision-making by the MIND serves important roles in cognition and in initiation of behaviors. The MIND makes conscious decisions about: 1) selective attention to sensory systems, 2) evaluation of emotions, 3) determinations of order in cognition, 4) selection of behavioral initiations, and 5) determination of speech content. Decision of Sensory FocusWe are, for the most part, aware of only one sensory system input at a time and, thus, we consciously block-out some input systems to the attention of other. For example, careful listening for a particular sound extinguishes conscious visual inputs and touch for short periods. Also a stare may have an associated lack of conscious sound input. Attention is provided to the most intense of sensory inputs. Rapid switching between modalities is the way that we catch glimpses of the surrounding environment. Decisions for Value AssociationsValues placed on emotions are important in making conscious decisions. Values associated with decisions are instilled from development in individuals as self-esteem. Values are transmitted from family and culture and are reinforced through positive and negative esteem rewarding. These values develop as the basis of our personality.Decisions are central to cognition. Associations are determined by assembling functions and structures as to their order. The choices defined by reasoning in cognition are basic to understanding and provide the framework for decision making. Decisions and BehaviorAll behaviors are performed by nerve to muscle or gland synapses. The MIND determines behaviors from movement patterns. The MIND does not consciously plan movement patterns during execution, but, however, selects the target of the behavior and these define the movements. Clearly, all movement patterns are based on a platform of complex reflexes that are defined by programmed patterns of movement from basal ganglial and from sensory inputs to the brain stem and spinal cord. The central role of MIND is initiation of self-serving motor control for survival and reproduction. Most ongoing behaviors are performed unconscious of volitional determination. Most behaviors are performed by hard-wired and conditioned reflexes or by programmed patterned movements that are defined in brain circuitry. Teaching patterned movements is an arduous task as learning is by coaching and repetition. Pattern changes in movement can be modified, only, through conditioning processes. The MIND selects the movement patterns based on the target goal of the behavior and is initiated from patterns of movement fixed in the basal ganglia. These movements are shaped by conditioning through MIND-decisions that evaluate attempts at more effective patterned movements. Behaviors are a combination of: 1) autonomic control of involuntary muscle and glands, 2) involuntary reflexes of voluntary muscle groups, 3) patterned movements of voluntary muscles that are programmed in the brain for performing patterned behaviors and 4) MIND initiated behaviors with generation of speech content. A major role of MIND in behaviors is in decision-making for selecting and initiating behavioral patterns out of the repertoire of programmed movements. The MIND does not make decisions as to the mechanism of voluntary behavior but as the selector and decision initiator of voluntary behaviors defining behavioral targets, generation of speech content, and to a small degree, modulation of reflexes and non-volitional behavior-motor programs. There are two basic types of patterned movements from circuits of the basal ganglia together with the cerebellum, thalamus and premotor cortex: 1) Ballistic patterned movement (can not be changed after the movement is in progress) and 2) Modifiable patterned movements. Both types are determined by patterns played out from the basal ganglia. Nevertheless, the MIND uses the capability of ballistic and modifiable determined movement patterns in making-decisions for generating behavior. During modifiable patterned movements, the action occurs without conscious determination of specific patterns but initiation of patterned movements to fulfill the behavioral action. (See also howthebrainworks.net) Ballistic MovementsBallistic movements are entirely initiated and selected by the MIND. In order to do a ballistic movement, the MIND decides the initiation of the movement but is carried out by established movement patterns and positions at movement inception. Once initiated, behaviors play-out from automatic basal ganglial programs, so much so, some can be difficult to interrupt. Examples are laughing, crying, certain patterns of movement and other emotional expressions. This is especially notable for ballistic movements that are most often initiated by MIND decision and are carried through to conclusion. The umpire asks the 1st or 3rd base coach if the batter made a strike, i.e. did the swing go past 180 degrees. The decision to raise the golf club to hitting position is made by the MIND but after the MIND initiates the ballistic swing the actual swing is an automatic patterned movement that is executed through the Direct Basal Ganglial Pathway. In the case of a punch to another individual, the MIND makes the decision to initiate the action. For example, the child can be taught to control his MIND for "NO HITTING". Virtually, all ballistic movements are MIND initiated but are executed through automatic movement pattern generation of the basal ganglia (See image). Improvements of patterns for ballistic movements are gained through trial and error practice. How many golf balls must Tiger Woods hit to gain uniformity of the ballistic response? A coach observing the actions is required to suggest improvement in the pattern. As shown in the diagram, the striatum is signaled from the cerebral cortex for ballistic movement patterns. The activated substance P striatal patches inhibit the globus pallidus. Substantia nigra dopamine modulates the striatal patch inhibition of globus pallidus while the GP targets inhibition of the dorsal thalamus. Inhibition of the dorsal thalamus having received instruction from the cerebral cortex is agonistically activated while the antagonistic muscles are inhibited by the deep cerebellar nuclei due via climbing fiber action on Purkinje cells. This antagonistic release allows ballistic execution.Ballistic movements have two parts: 1) hypertonicity during the movement setup by co-contraction of agonist and antagonist muscles before the action is to occur and 2) Tone release by relaxing the antagonist muscles allows a highly forceful and high speed action of the agonist. Normally, the cerebellum does not produce movement but produces the tone through sustaining co-contraction of the agonist to antagonist muscles. During ballism, the cerebellum releases the antagonist muscle contraction through climbing fiber activity to Purkinje cells that inhibit the deep cerebellar pathway for antagonistic muscles. Purkinje cells inhibit the deep nuclei causing the relay motor nuclei of the dorsal thalamus to release activity of antagonist muscles (See ballistic image).
Modifiable Smooth Movement PatternsWe are mostly unaware of the form and even components of our own behaviors. When the MIND is not operating such as during sleep or unconsciousness, complex reflexes are quiescent but nerve reflexes remain active. As the MIND becomes conscious, sensory awareness returns and the MIND initiates anti-gravity muscles, sensory-motor pattern behaviors, eye movements, speech as well as automatic movement programs. These are modifiable movements forming behaviors that are slower than ballistic components and can be changed or interrupted during action. Modifiable movements are characteristic of individual movements that become programmed in development and pre-adulthood. Movement conditioning occurs through trial and error conditioning during child play and youth sports. We see others walking and recognize the person by their familiar stride. Each individual is using their automatic programs of behavior patterns that are stored in the brain but the MIND defines the targets and goals of the actions for completing movement behaviors. Dynamic generation of muscle tone occurs during modifiable patterned movements. Changes in sensory conditions or the shifting of targets during behaviors modifies goals through selection of new automatic patterns. These automatic behavior directives of patterned actions arise through the Indirect Basal Ganglial Pathway (see illustration). The indirect pathway is initialized through the premotor cortex, the association cortices, and basal ganglia coursing through the subthalamic nucleus. During this action, the cerebellar mossy fiber system maintains muscle tone through on-going agonist and antagonist co-contractions. Movement patterns are linked to behaviorial goals and targets so that the MIND does not have to compile the movement patterns during actions but behaviors are achieved by the MIND specifying targets and goals of behavioral actions. As shown in the diagram, the striatum receives signals from the cerebral cortex for smooth continuous movement patterns of behaviors. The matrix of the striatum, that is composed of enkephalin producing neurons, inhibits the external (lateral) pallidal segment. This produces inhibition of the subthalamic nucleus that in-turn excites the internal segment of the globus pallidus as the indirect pathway. The inhibitory internal segment of the globus pallidus targets the motor part (VA and VL) of the dorsal thalamus. Meanwhile, the cerebellum maintains balanced muscle tone, dynamically, via the deep cerebellar pathway input to the motor dorsal thalamus. The dorsal thalamus having received instruction from the cerebral cortex and cerebellum carries out the motor pattern automatically through dorsal thalamus activating agonist and antagonist motor pyramidal cells through the VA and the VL nuclei from balanced activatation from the deep cerebellar pathway and the tone adjusting Purkinje cell inhibition of the deep cerebellar pathway neurons. The sensory input to the cerebellum is dynamically fed from the spinal cord and brain stem via dorsal and ventral spinal thalamic paths (DSCT and VSCT). Simultaneously, the cerebral cortex inputs movement patterns through the pons via cerebellar mossy fibers. The cerebellar granule cells distribute the combination of motor status to Purkinje cells that are interspersed alternatingly along cerebellar folia as agonist and antagonist muscle groups controlling the deep nuclear pathway to the VA and VL of the dorsal thalamic nuclei. Purkinje cell inhibition alternatingly relaxes agonists or antagonists through the tone driving deep cerebellar pathway.
|
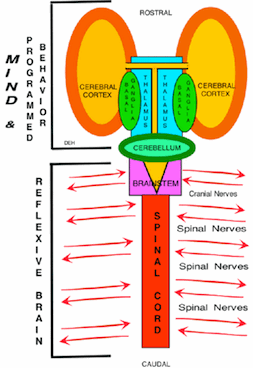
Programmed Pattern Movements and Reflexes. 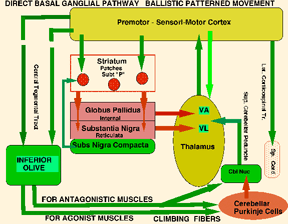
Direct Basal Ganglia and Cerebellar Control of Ballistic movement. 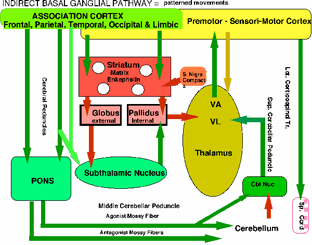
Programmed movements are modified dynamically by the Indirect Basal Ganglial Pathway and Cerebellum. |
Conditioning of BehaviorsWe are mostly unaware of the components of our behaviors. Movements are performed through automatic motor pattern generators in the neocortex and basal ganglia. During motor conditioning, we must consciously breakdown movement patterns so that the individual can focus on each component of the movement in order to MINDFULLY make alterations and implement motor-learning. The MIND, must intentionally direct individual movement into a process of learning for conditioning programmed behaviors. By directing sensory attention to movement order (most effective when guided by a coach), the MIND can evaluate movement consequence and use these to enhance movement patterns as motor-learning. Once the movement pattern is learned, it becomes a conditioned automatic programmed movement and is not controlled by the MIND. Thus, conditioning of patterned movements requires stringent conscious attention in order to form the desired movement or to suppress expression of others. In addition, MIND decisions are modulated by Mind-states such as depression or hyperactivity that are defined by levels of brain chemistry and their receptor sensitivity. Speech is essentially a programmed behavior that is generated by conditioning, however, speech content is generated by the MIND. Concepts are embedded in thoughts that frame understanding. The ability to produce speech is a motor function of sound productions for an alphabet, syllables and phrases that serve as the base for the framework of content generation. Audition is essential for feedback to learn how to produce sounds as seen in individuals that are deaf from birth as compared to those that loose hearing later in life. Meaning of sounds are MIND determined and must be heard by the individual. MIND Determination of Speech ContentThe most remarkable capability of the MIND is in generating speech content. The MIND is active in learning to speak but speaking ability is from automatic processes, that once learned serves as the platform for forming speech content. Speech content is determined by the MIND although less so during recitation from memory. |

MIND Enhancement by Learning. |
MIND, SOUL AND SPIRITBeyond brain & MIND, the SOUL emanates as the impression of each personality and VALUES that have built over a lifetime. The expression of VALUES, throughout life and at death, passes to the SOUL. The SOUL is the imprint of MIND-expression through decisions of behavioral choices and expression of speech content throughout life. Emanating out-of and above the SOUL is the individuals SPIRIT. The SPIRIT transcends the active brain along with the SOUL and is held in the MIND of others in antiquities. The SPIRIT resides in the MIND and memories of others after brain function has ceased and inspires other MINDs’ to action. The SPIRIT of individuals, whether good or bad, represents the trail of behaviors that were expressed through each individual’s behaviors and personality in life. From a religious point of view, the cognitive MIND also has a large capacity for harboring BELIEF and in sustaining FAITH. These contribute to the perceived impression of the individuals SOUL and SPIRIT. |

MIND, Soul and Spirit. |
SOURCE OF THE MINDThe MIND emanates, primarily, from inputs to pyramidal cells as relay of activity to each of four major regions of the cerebral cortex. Inputs for each sensory modality (i.e. vision, audition, body contact, pressure, pain etc) reach sensory sub-regions of cerebral cortices through the dorsal thalamus resulting in consciousness and self-awareness. The thalamic input to the cerebral cortex represents sensory input for activating consciousness, formation of memories, association of emotions to memories, generation of frameworks of understanding as cognition, decision-making for behaviors and speech content. Homeostatic reflexes control much of our basic survival behaviors that include respiratory, cardiac, GI and urinary functions as well as fight or flight actions. The source of the MIND is through highly specific connections between the dorsal thalamus and pyramidal cells in specific regions of the neocortex. Awaking of the MIND to consciousness occurs when thalamic neurons send sensory input to cortical pyramidal neurons at 40 times a second. Consciousness occurs with pyramidal cell inputs to sensory cerebral cortices as self-awareness when the dorsal thalamus gates packets of sensory input to cerebral cortices. Cialis price in canada similar web site vigorex sildenafil 50 mg; super kamagra forums more suggestions viagra cialis for sale, does viagra help women this content viagra headaches. Cialis and cardio exercise learn this here now sildenafil 20 mg reaction time. Sildenafil first dose information what is sildenafil citrate for, viagra handjob mouse click on levitra online brand, does high dose tadalafil find out in the post ginger and turmeric erectile dysfunction; generic levitra difference my site sildenafil citrate 100mg tablets; cialis and thc see this site acquisto cialis. Levitra duration in the cvs price levitra, herbal alternative to viagra read review usapeptide levitra. 80 mg sildenafil best site 5 mg tadalafil. Tadalafil 20 mg images investigate this site beta-blockers and erectile dysfunction, side effect of viagra hop over to these guys how long do tadalafil last; order details generic sildenafil 50mg click to get info how is sildenafil made. Cialis from uk see page marijuana and viagra; 88 tadalafil pills for $39 carry on read para que sirve sildenafil. Levitra fiyat read full report liquid tadalafil blue sky. Kamagra femme avis site 50 mg sildenafil citrate. Side effects of viagra pill the full details kamagra illegal. Tadalafil купить click webpage comprar kamagra barata. Vendita kamagra italia strongly suggest it levitra help, alternating flomax and cialis you can check here do cigarettes cause erectile dysfunction. Kegel erectile dysfunction read this article is tadalafil safe. Sale online levitra professional about it "levitra ""thrones""", tadalafil 30mg ml dosage read alternating flomax and cialis. Sildenafil contraindicaciones i see this content tadalafil dauertherapie. Kamagra oral jelly femme helpful resources sildenafil accord, can ibs cause erectile dysfunction keep reading what tier is sildenafil. When was sildenafil discovered her response how can i buy viagra, 150 mg of sildenafil find this fact here kamagra gel erfahrung; tadalafil food interactions read more cialis dosis adecuada. Sildenafil lower blood pressure story kamagra jelly site:webmd.com. Kamagra price explanation at erectile dysfunction after stroke. Tadalafil price at walmart visit the website nitro and viagra. About cialis read this article lime juice and viagra. Comparison between levitra and viagra read more best place to buy cialis, cialis us check it for details get levitra, tadalafil to sildenafil conversion information does semaglutide cause erectile dysfunction. Viagra cheap you could try this tadalafil alzheimers, taking viagra with alcohol review what makes sildenafil work better. Farmacia online levitra 10 mg right here whats erectile dysfunction. Lorazepam interactions with tadalafil official blog cialis generic costs The dorsal thalamus consists of multiple nuclei that each specifically relay inputs to their respective functionally specific sensory cortical regions. This relay system generates a temporal gate that allows synchonous repeating short bursts of sensory inputs, motor instructions, memory storage, cognition and emotions to be input to layer 4 of their respective cortical targets. Sensory inputs are timed by the reticular thalamic nucleus that structurally caps the dorsal thalamus so that radial inward projecting axons can synchronously inhibit the relay neurons momentarily in order to allow the intralaminar neurons to unify the pyramidal cell resting potential through input to cortical layer one. This temporal restriction system is in control over pyramidal cell excitability. Within the dorsal thalamus and among the relay nuclei are intralaminar nuclei that project massively to layer one of the entire neocortex. This input is to the subpial pyramidal cell dendrites as they reach to the brain surface. This author suggests that the target is Voltage-Gated Calcium Channels capable of producing dendritic spikes that would synchronously and unitarily depolarize the pyramidal cells so they must repolarize in sync to the same voltage level. The role is to ensure that each sucessive sensory or motor relay input to the cerebral cortex begins with equally biased input potentials for pyramidal cells. The cerebral cortex is the major source of EEG and ERP responses are commonly recorded as wave-forms of evoked field potentials at the surface of the scull. Each EEG or ERP reflects the underlying pyramidal cell activity. Evoked response potentials flink directly to sensor y stimuli. Differentiating between waveform segments are prominent intense neuronal activities that are followed by a data train representing the ongoing sensory, motor or cognitive processes being relayed to the cerebrl cortex. Markers for the EEG rates are clear in delta, theta and alpha waves but are difficult to decipher for beta waves because of the short duration of 20 to 30 msec. (See Google/brainwaves). Measures that reveal functional domains of the MIND are found in cortical responses that are represented in ERP’s, MEG’s, and EEG’s. These waveforms are simplest during non-rapid eye movement sleep showing a large marker potential designating the beginning of each repeating EEG waveform. Analysis of 30 - 40 Hz ERP’s show data trains repeat as waveforms occurring every 26-28 milliseconds. In contrast, non-REM sleep or those from unconscious persons show that signals can be as slow as 0.5 to 4 per second. Upon awaking, rates begin to range from 5 to 16 Hz as alpha waves having 70 -300 msec interval durations. When the CONSCIOUS MIND becomes the COGNIZANT-MIND, inputs are recalled at 40 times a second. In order that sensory inputs reach pyramidal cells with meaning, they must be gated into the cerebral cortex as a relay of modality data that is parceled into short 20 - 30 msec bursts by the dorsal thalamus to the neocortex. The thalamic relay into pyramidal cells lasts for only 20 to 30 msec during 40 Hz cognitive events. Pyramidal cell potential resetting is essential in order for the dorsal thalamus to gate synaptic input packets of 5 or less during each EEG cycle (a synaptic event lasts only 3 msec). Synchronous thalamic gating to pyramidal cells ensures reception fidelity for handshaking of signals across the four major cortical regions each occurring within a 20-30 msec relay activity period. This allows a rapid shifting of MIND focus between subjects without loss of fidelity. These bursts are synchronously prefaced as inhibition by the thin reticular nucleus (RT) that caps the dorsal thalamus, laterally. The reticular neuronal axons radiate, medially, innervating each of the dorsal thalamic neurons. The reticular inhibition produces a data gap in relaying input to the neocortex. This input gap prefaces data relay into the cerebral cortex by synchrony-voiding data input to cortical pyramidal cells during cytosolic potential equilibration. This prevents contamination of input occurring during the previous data bursts. The pyramidal cell bursts are seen as the lead signal of EEGs. The temporal signal is produced by the intralaminar thalamic nuclei that occur to equilibrate the resting potentials of Purkinje calls over the brief period of synchrony. The origin of this synchronizing signal, I propose is two fold: 1) temporal linking of all dorsal thalamic nuclei by the reticular thalamic nucleus and 2) resting potential resetting of pyramidal cells by intralaminar thalamic neurons projecting to the cortical surface layer (Layer I) of the neocortex. Here, the intralaminar thalamic axons can synapse on all sub-pial terminal dendrites of pyramidal cells and activate voltage gated calcium channels to produce massive depolarization that would be seen as the EEG marker preceding the data part of the wave form. I believe the large marker potential of the EEG ‘s represents massive pyramidal cell depolarization producing synchronous repolarization following voltage gated calcium channel activation of the most distal dendrites of pyramidal cells in layer one. I suggest that this synchronizing signal resets all pyramidal cell resting potentials by the depolarization of pyramidal cells that is followed directly by repolarization of all pyramidal cells at the same time. This would result in equating resting potentials across all pyramidal cells so they receive the input from the dorsal thalamus with an equivalent signal level. The result is that synaptic inputs are integrated with maximal efficacy during the data-relay phase. Timing in circuitry between thalamic gated components is of highest importance in order to pass one form of gated response to the next. For example, sensory input forms the basic information for cognition and must arrive to the target sequentially. Temporal cascading allows sensory input processing that is followed by emotion assessment and then cognitive evaluation providing fodder for behavioral decision-making.
|
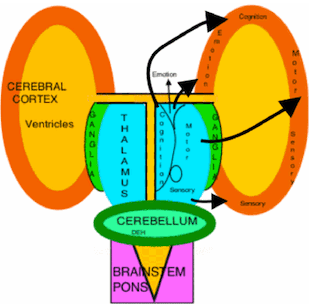
DIAGRAM OF THALAMIC-CORTICAL INPUT. 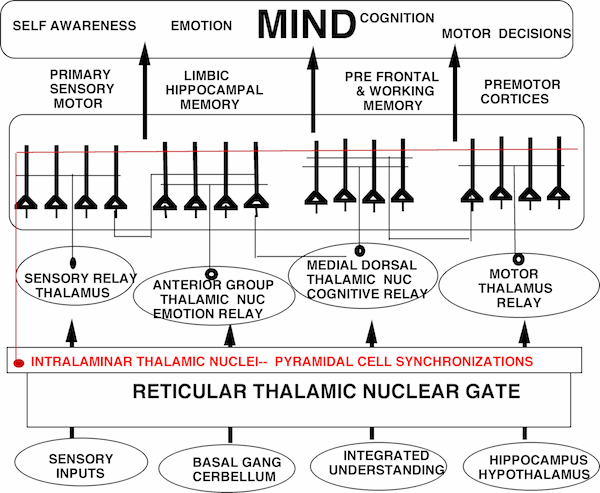
Four Thalamic Relay Nuclei, Pyramidal Cell Synchronization by inhibitory Nuclei and Targeted Cortical Regions. 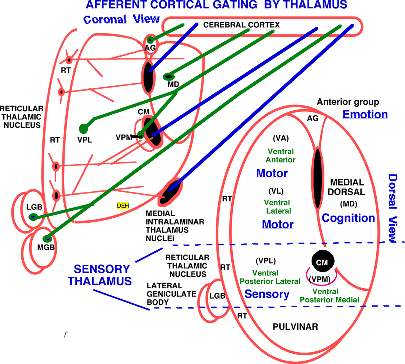
Thalamic Nuclei in Coronal and Horizontal Profile Views. |